
Research progress on the role of G-protein-coupled receptors in T lymphocyte immunomodulation
-
摘要: G蛋白偶联受体(GPCRs)是一类跨膜受体超家族,参与多种信号转导通路,在细胞迁移、代谢等生理过程中发挥重要功能。T淋巴细胞是重要的免疫细胞,参与炎症反应过程并在细胞免疫中发挥重要作用。目前已发现多种GPCRs在T淋巴细胞中表达并参与T淋巴细胞免疫调节过程,本文将对GPCRs在T淋巴细胞免疫调节中的作用进行综述。Abstract: G-protein-coupled receptors (GPCRs) are a type of superfamily of transmembrane receptors, involved in multiple signaling pathways, and playing an important role in physiological processes such as cell migration and metabolism. T lymphocytes are important immune cells that participate in the inflammatory process and play an important role cellular immunity. To date, multiple GPCRs have been found to be expressed in T lymphocytes and participate in T cell immunomodulatory processes. This paper reviews the role of GPCRs in T lymphocyte immunomodulation.
-
Keywords:
- G-protein-coupled receptors /
- T lymphocytes /
- immunity /
- tuberculosis
-
G蛋白偶联受体(G-protein-coupled receptors, GPCRs)是一类膜蛋白受体的总称,是人体内最大的膜受体蛋白家族。GPCRs可结合多种细胞外信号,通过激活G蛋白将细胞外信号传递到细胞内部,在调节机体生理功能中起到重要作用,并在其发挥作用的过程中受到多种分子的调控[1-4]。GPCRs是药物研发的重要靶点之一,尤其是对心血管、神经、免疫等相关疾病的药物研发。目前靶向GPCRs的药物约500个,占美国食品药品监督管理局(U.S. Food and Drug Administration, FDA)已批准药物的34%,占全球市场的27%[5-9]。
T淋巴细胞(简称T细胞)在细胞免疫中起重要作用,T细胞功能紊乱可引起多种疾病,如感染、过敏等。目前在T细胞表面已经发现了多种GPCRs,其中许多都参与了T细胞活化、迁移和细胞周期调控并发挥关键作用。
T细胞来源于骨髓的淋巴干细胞,在胸腺中分化、发育成熟,通过淋巴和血液循环分布于全身的免疫器官和组织。受到刺激后,细胞外信号分子通过多种信号通路介导效应T细胞活化,活化后的CD4+T细胞分化为不同的效应T细胞亚群,产生多种细胞因子。研究[10-11]发现,CD4+T细胞亚群Th1和Th2可释放白细胞介素(interleukin, IL)-1、IL-2、IL-4、IL-5、IL-6、肿瘤坏死因子(tumor necrosis factor, TNF)-α、TNF-β、干扰素γ等多种炎性因子促进T细胞介导的炎症反应。活化后的CD8+T细胞通过穿孔蛋白和Fas配体(Fas ligand, FasL)杀伤靶细胞。研究[12]发现,GPCRs及其信号通路在T细胞免疫过程中起着重要作用,如参与T细胞的激活,维持细胞稳态,参与细胞代谢。
1. GPCRs信号
GPCRs包含7个跨膜蛋白受体超家族,可与离子、神经递质和趋化因子等多种细胞外配体进行结合,并通过与三磷酸鸟苷(guanosine triphosphate, GTP)相互作用进一步激活细胞膜上的G蛋白。G蛋白由α、β和γ三个亚基组成,根据α亚基的不同,G蛋白进一步分为四个不同的亚家族:Gs, Gi/o, Gq/11和G12/13。在静息状态下,α亚基及β、γ亚基与二磷酸鸟苷(guanosine diphosphate, GDP)结合。当GPCRs被激活后,GTP结合G蛋白的α亚基,引起α亚基与β、γ亚基分离,与下游信号分子相互作用,调节细胞功能[13]。GPCRs与Gs结合后激活腺苷酸环化酶,升高细胞内环磷酸腺苷(cyclic adenosine monophosphate, cAMP)水平。GPCRs与Gi结合可抑制腺苷酸环化酶活性,细胞内cAMP水平降低。GPCRs与Gs结合后可激活磷脂酶C,磷脂酶C可以将磷脂酰肌醇二磷酸[phosphatidylinositol(4, 5)bisphosphate, PIP2]转化为二酰基甘油酯(diacylglycerol, DAG),进而激活蛋白激酶C(protein kinase C, PKC),介导钙离子释放。同时,GPCRs信号还可以受到G蛋白信号传导调节蛋白(regulators of G protein signaling, RGS proteins)、GPCRs激酶(G protein-coupled receptor kinases, GRKs)和β-阻滞蛋白(β-arrestin)等调节[14]。在T细胞表面已经发现了多种GPCRs,下面将列出一些与T细胞功能与调节相关的典型GPCRs。
2. GPCRs可调节T细胞迁移
2.1 C-C趋化因子受体(chemokine receptor, CCR)5
CCR5是GPCRs家族中的一种细胞膜蛋白,可在T细胞、巨噬细胞及小胶质细胞表面表达,结合CCL3、CCL4及CCL5,是T细胞活化和动员的重要调节剂,可参与T细胞向感染部位的迁移[15-17]。Hoft等[16]研究表明,CCR5与配体结合后可促进Th1细胞迁移至小鼠的结核分枝杆菌感染的部位。Harlin等[18]发现,小鼠黑色素瘤部位CD8+T细胞CCR5表达水平升高;当缺乏CCR5配体时,黑色素瘤小鼠体内T细胞在肿瘤部位浸润减少。Tan等[19]发现调节性T细胞(regulatory T cells, Treg)可迁移到肿瘤微环境,这一过程是由CCL5-CCR5轴介导,当CCR5抑制剂存在时,Treg细胞的迁移减少。Tavares等[20]研究表明小鼠感染甲型流感病毒后,体内释放炎症因子,信号分子通过CCR5通路诱导T细胞募集到感染部位,进而形成支气管相关淋巴组织(inducible bronchus-associated lymphoid tissue, iBALT),减少体内病毒量并改善肺功能;而CCR5敲除小鼠体内的T细胞募集数量较少,肺损伤及肺功能障碍程度较重,导致感染流感病毒后的病死率较高。CCR5还参与呼吸道病毒感染期间记忆性CD8+T细胞向肺气道的迁移[21]。此外,CCR5可与CXCR4相互作用,调节T细胞的活化[22]。
2.2 GPR56
GPR56是一种黏附型GPCRs(adhesion G protein-coupled receptors, aGPCRs),细胞外N端结构域包含较多的丝氨酸和苏氨酸残基以及GPCRs蛋白水解位点(GPCRs proteolysis site, GPS)结构域,这些结构在介导细胞功能时起到重要作用。Peng等[23]研究发现,在巨细胞病毒感染期间,体内效应CD8+T细胞稳定表达GPR56,并且GPR56与细胞毒性T细胞(cytotoxic T lymphocyte, CTL)的迁移有关,与对照组相比,GPR56表达上调可抑制T细胞迁移。此外,Liu等[24]研究表明,GPR56信号通路与CTL发挥毒性功能有关,GPR56可成为鉴别T细胞的标志物。
2.3 趋化因子受体(CXCR)3
CXCR3是GPCRs家族中的一员,可在Th1细胞及CTL细胞表面表达,与趋化因子配体(CXCL)9、CXCL10和CXCL11等结合。CXCR3与配体结合后可引起多种细胞反应,在调节T细胞迁移过程中发挥重要作用[25-26]。Li等[27]发现,与野生型小鼠相比,CXCR3-/-小鼠在肝移植切口处Tregs细胞的动员和募集较少,而CXCL10/CXCR3信号通路激活可促进肝移植切口处Tregs的动员。Kuo等[28]发现CXCR3信号通路介导的T细胞募集可导致皮肤移植排斥,CXCR3与配体结合后促进T细胞的浸润,使供体发生皮肤移植排斥反应。CXCR3除了参与T细胞募集与迁移外,还参与T细胞的分化。Karin等[29]发现,CXCL10与CXCR3结合后,可介导STAT1、STAT4和STAT5等磷酸化,STAT1与STAT6磷酸化后可引起T细胞分化为Th1细胞。CXCL11与CXCR3结合后,通过mTOR通路参与CD4+T细胞分化,形成分泌IL-10的Tr1。此外,有研究[30]发现CXCR3缺陷的CD8+T终末分化水平较低。
2.4 CCR4
CCR4是GPCRs家族的一员,CCR4的配体包括CCL17和CCL22[31-32],CCR4在T细胞的迁移中起着重要作用。一项小鼠相关研究[12]发现,树突状细胞活化后可激活T细胞,促进T细胞表达CCR4,与对照组相比,被激活的CCR4缺陷T细胞迁移至肺部的数量明显减少,树突状细胞可通过诱导T细胞表面CCR4表达,进而促进T细胞迁移至肺。调控Th17细胞的CCR4/TARC信号轴可介导T细胞向肺内的迁移[33]。因此,利用CCR4抗体阻断趋化因子与CCR4的结合,抑制Th2细胞向炎症部位迁移,是控制变态反应的新靶点。CCR4在肿瘤患者体内T细胞表面高表达[34],CCR4的高表达与肿瘤性疾病的高风险进展相关[35],并且CCR4+T细胞的数量在皮肤T细胞淋巴瘤的病情后期扩散时明显增加[36]。CCR4还与多种过敏性炎症疾病有关,如哮喘、特应性皮炎和变应性鼻炎[37],Honjo等[38]研究表明,在过敏性肺炎的小鼠模型中,CCR4为抗原特异性Th2细胞进入肺和气道所必需,机体处于炎症感染时,CCR4+CD4+T细胞的聚集可被CCR4抗体K327明显抑制,且其抑制作用与剂量呈正相关,因此,CCR4可作为临床治疗AR的新靶点[39]。
2.5 CCR7
CCR7属于GPCRs家族,可在T细胞表面表达,介导T细胞迁移[40]。配体CCL19、CCL21与CCR7结合后可介导细胞外信号传导[41-43]。在小鼠试验中,CCR7是CD4+T细胞迁移至淋巴组织的重要受体[44],敲除CCR7后的T细胞不能迁移至淋巴结[45]。此外,在适应性免疫中,次级淋巴组织(secondary lymphoid tissues, SLTs)内CCR7介导的T细胞迁移发挥重要作用[46]。含有SH2结构的5’肌醇磷酸酶-1(SH2-containing inositol phosphatase-1, SHIP-1)表达上调可促进CD4+T细胞表面CCR7表达,进而促进CCR7介导的T细胞向局部淋巴结的转移,拮抗细胞SHIP-1表达可抑制此迁移过程[47],表明CCR7在T细胞迁移过程中发挥重要作用。CCR7在免疫系统疾病中也发挥重要作用。原发性干燥综合征(primary Sjogren’s syndrome, pSS)患者体内T细胞表达CCR7明显上调,加入CCR7抗体后,pSS患者的CD4+T细胞迁移能力明显下降,表明CCR7是T细胞迁移的重要因素。T细胞表面的CCL21/CCR7被活化后可激活细胞内JNK和p38MAPK通路,阻断两条通路后,pSS患者体内CD4+T细胞的迁移明显减少,提示JNK和p38MAPK通路在pSS患者体内CD4+T细胞的迁移过程中发挥重要作用,是治疗pSS的新靶点。免疫组化结果显示,在皮肌炎(dermatomyositis, DM)患者的炎症浸润部位,CCR7表达明显升高[48]。此外,在DM患者的肌肉组织匀浆中,CCR7的表达显著上调,表明CCR7可能参与了DM患者炎性T细胞的募集[49]。
3. GPCRs可调节T细胞分化
3.1 嘌呤受体P2Y6(purinergic receptor P2Y, G protein-coupled, 6)受体
P2Y6受体属于GPCRs中的P2受体家族,可被细胞外核苷酸激活,在调节T细胞对炎症的反应中起着重要作用,P2Y6受体参与包括气道炎症反应在内的多种炎症反应[50]。研究[51-52]表明,在细菌和病毒诱导的机体免疫反应中,P2Y6受体发挥重要作用。此外,P2Y6受体在T细胞活化中发挥作用,P2Y6表达可抑制效应T细胞的活化[53-55]。
3.2 G2A
G2A是GPCRs家族的一员,可在淋巴细胞和巨噬细胞表面表达。G2A的配体包括溶血磷脂酰胆碱(lyso-phosphatidylcholine, LPC)。LPC是一种促炎脂质,可激活G2A,诱导T细胞募集到炎症部位[56-57],此外,有研究[56-60]发现,LPC通过G2A可激活ERK1/2、JNK、MAPK等下游信号分子,进而通过升高TGF-β1表达水平促进Tregs功能,其中JNK是LPC诱导nTregs中TGF-β1表达的主要因素。G2A与LPC结合后能够调节T细胞迁移,还参与自身抗原特异性T细胞的迁移,介导慢性自身免疫性疾病的发展;另一方面,G2A可负责调节T细胞对抗原的特异性反应,通过减少自身抗原特异性T细胞的产生来减弱免疫反应。G2A在T细胞自身免疫性疾病中的作用还需要进一步研究。
3.3 EP
EP是位于细胞膜的GPCRs家族成员之一,可与前列腺素E2(prostaglandin E2, PGE2)结合。EP分为EP1、EP2、EP3和EP4四种不同亚型,参与不同的信号传导途径,调节免疫反应,发挥生理功能[61]。EP1与Gq结合后可被激活,升高Treg细胞内PGE蛋白及钙水平,激活PKC,进而激活NFAT/NFκB/MAPK信号通路,诱导基因转录,发挥生理功能[62]。EP受体可调节T细胞介导的免疫反应。Li等[62]发现PGE结合EP2后可通过cAMP通路降低Foxp3 mRNA表达水平,进而抑制T细胞分化。Nataraj等[63]发现激活EP1可增强Th1介导的免疫反应。Boniface等[64]发现PGE结合EP2、EP4后可通过cAMP通路活化Th17细胞。Sreeramkumar等[65]发现EP4在Th1细胞活化过程中表达水平明显升高,抑制CD4+T细胞表面的EP受体会减少炎症期间CD4+T细胞向炎症部位迁移,减少CD4+T细胞的增殖。
3.4 G蛋白偶联受体激酶(GRK)2
GRK2属于GPCRs家族成员,在T细胞表面表达,并在介导内吞、细胞内运输、机体脱敏等过程中起重要作用[66]。研究[67]表明,GRK2可通过磷脂酰肌醇-3-激酶(phosphatidylinositol-3-kinase, PI3K)、AKT、丝裂原活化蛋白(mitogen-activated protein, MEK)等通路介导炎症、心血管和肿瘤等疾病的进展。此外,GRK2在T细胞中的表达与哮喘发病有关,有研究[68]发现GRK2可激活T细胞受体(T cell receptor, TCR)信号,促进IL-2和IL-10等细胞因子的产生。GRK2在哮喘患者体内和暴露于屋尘螨(HDM)的小鼠的肺内表达增强。在HDM诱导的小鼠过敏性哮喘模型中发现,与对照组相比,敲除T细胞表面GRK2受体的小鼠体内IL-4、IL-13水平降低,支气管肺泡灌洗液(BALF)淋巴细胞计数和黏液分泌明显减少,肺部炎症加重,提示GRK2在哮喘相关T细胞活化中发挥重要作用[69]。此外,GRK2可通过介导PI3K-AKT通路调节Tregs分化及发挥功能[70-71]。研究[68]表明GRK2参与TCR激活CXCR4,并介导CXCR4的磷酸化,进而促进TCR-CXCR4复合物形成。此外,TCR介导的Src激酶激活可能导致GRK2磷酸化,从而影响T细胞活化。
4. GPCRs可调节T细胞代谢
4.1 腺苷A2A受体(A2AR)
T细胞表面可表达A2AR。A2AR结合腺苷分子,参与细胞信号传导,是炎症和免疫应答的重要受体[72-73]。CD8+T细胞可通过A2AR信号传导通路抑制T细胞的效应功能,如抑制T细胞增殖、活化,并且抑制T细胞分泌促炎因子(IFNγ、TNFα)。A2AR可结合Gs蛋白,增加cAMP和蛋白激酶A(protein kinase A, PKA)水平,通过细胞内A2AR/PKA/mTORC1信号通路抑制T细胞活化,引起T细胞代谢障碍。此外,A2AR激动剂可减少CD8+T细胞IFN-γ的产生[74-77]。Mastelic-Gavillet等[78]发现,腺苷与A2AR结合后,可降低CD8+T细胞的糖酵解代谢活性,损伤CD8+T细胞活性,相反,当选择性抑制A2AR后,由腺苷引起的CD8+T细胞损伤减少。
4.2 GPR30
GPR30是GPCRs家族中的一员,分布于细胞膜及内质网,在多种组织和细胞中表达,调节机体骨骼发育、生殖等生理功能。GPR30结合雌激素后可升高细胞内Ca2+浓度,诱导细胞核合成磷脂酰肌醇3, 4, 5-三磷酸(phosphatidylinositol 3, 4, 5-trisphosphate, PIP3),参与细胞的存活、增殖过程[79-80]。Prossnitz等[81]发现,雌激素可通过结合GPR30调节CD4+T细胞的免疫功能,GPR30特异性激动剂G-1可增强T细胞的免疫抑制,促进细胞凋亡,抑制炎症因子的产生。Wang等[82]发现GPR30敲除小鼠可消除体内雌激素诱导的脾细胞CD4+FoxP3+Treg中程序性死亡受体(programmed cell death protein, PD)-1表达上调,在Treg细胞中雌激素通过GPR30和PD-1介导的机制增强Treg细胞的抑制活性。研究[83]发现,长期雌激素治疗会导致胸腺萎缩及胸腺细胞功能丧失,Wang等[84]发现雌激素可通过GPR30介导胸腺萎缩,在E2诱导的胸腺萎缩模型中,GPR30敲除小鼠的胸腺萎缩程度比对照组明显降低,GPR30激动剂可诱导胸腺萎缩和胸腺细胞凋亡。
5. GPCRs与感染
GPCRs在调节T细胞功能中起到重要作用,而T细胞作为免疫细胞的重要组成部分,在机体感染病原体过程中发挥作用。Fisher等[85]发现,恙虫病感染期间鼠脑组织的差异基因表达分析发现T细胞中CXCR3基因表达较对照组明显升高。GPCRs在结核分枝杆菌感染过程中也发挥重要作用。Mamtani等[86]发现,CCR5及其配体的促炎作用与活动性结核病发展密切相关。Shanmugasundaram等[87]发现结核分枝杆菌潜伏感染者气道内的大部分特异性CD4+T细胞表达CXCR3,且患者肺部的CXCR3+CD4+T细胞数量与结核分枝杆菌浓度呈负相关。Park等[88]发现,经结核分枝杆菌的RpfE蛋白刺激后的树突状细胞可产生PGE2,产生的PGE2主要通过EP4受体诱导Th1和Th17细胞分化,进而发挥较强的抗结核分枝杆菌活性。GPCRs也参与病毒感染后引起的免疫反应。Kuo等[25]发现,当单纯疱疹病毒感染皮肤后,CD8+T细胞表面CXCR3表达上调,且CXCR3的配体CXCL9、CXCL10水平高于对照组皮肤细胞,CXCR3与配体结合后可促进CD8+T细胞募集到感染部位,有助于清除病毒。当皮肤感染牛痘病毒时,表达CXCR3的CD8+T细胞迁移至牛痘病毒感染部位,发挥细胞杀伤作用。此外,CCR1可调节EB病毒(Epstein-Barr virus, EBV)基因表达水平、癌细胞播散和T细胞迁移[89]。因此,在自然杀伤/T细胞淋巴瘤(natural killer/T-cell lymphoma, NKTCL)中,靶向CCR1的治疗有助于EBV清除、T细胞活化和淋巴瘤细胞杀伤,是治疗EBV诱导癌症的新方向。GPCRs在人类免疫缺陷病毒(human immunodeficiency virus, HIV)感染机体过程中也发挥作用。CCR5和CXCR4能够直接与HIV表面糖蛋白GPl20结合,促进HIV-1病毒进入T细胞,是HIV-1和HIV-2病毒入侵细胞的关键步骤。配体CCL5、CXCL12分别与CCR5、CXCR4结合后可抑制HIV-1感染机体过程[90]。目前,使用CXCR4和CCR5拮抗剂治疗HIV在临床研究方面已取得一定成果。组织驻留记忆(tissue-resident memory, TRM)细胞促炎活性较强,在急性感染期间,TGF-β诱导GPR56在TRM细胞表达上调,抑制TRM细胞的活性,避免炎症引起的机体损伤。此外,在淋巴细胞性脉络丛脑膜炎病毒(lymphocytic choriomeningitis virus, LCMV)和李斯特菌急性感染时,GPR56在CD8+TRM细胞中特异性上调[91],调节CD8+T细胞的分化及功能。在流行性感冒病毒感染期间,气道炎性上皮分泌的CXCL9和CXCL10增加,募集表达CXCR3趋化因子受体的CD8+T细胞[92]。综上所述,GPCRs在病毒感染过程中发挥重要作用,探究在病毒感染过程起关键作用的GPCRs可为病毒药物的研发提供新的靶点[93]。
T细胞中的GPCRs的功能见图 1,GPCRs在T细胞中的作用见图 2。
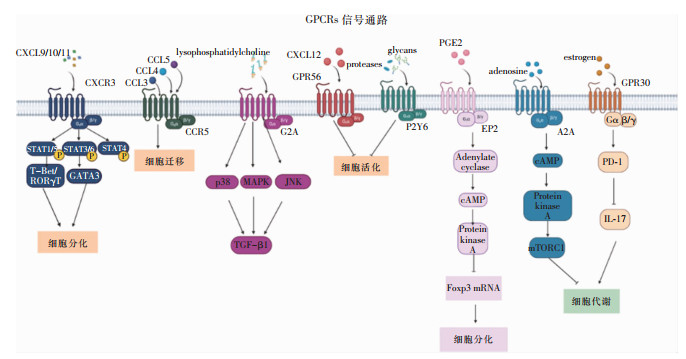 图 1 T细胞中GPCRs的功能
图 1 T细胞中GPCRs的功能 下载:
全尺寸图片
下载:
全尺寸图片
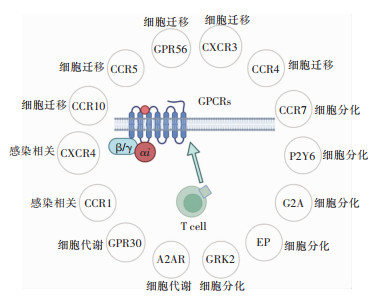 图 2 GPCRs在T细胞中的作用下载:
全尺寸图片
图 2 GPCRs在T细胞中的作用下载:
全尺寸图片
6. 总结与展望
综上所述,T细胞参与多种适应性免疫反应,T细胞功能紊乱与自身免疫性疾病有关。GPCRs及其调控分子在T细胞生物学功能中起着重要作用。本文列出了一些参与调节T细胞功能的GPCRs信号通路及其调控因子,以及在T细胞发挥功能的过程中GPCRs介导的信号通路成分。未来对这些分子的研究将会给T细胞生物学提供新视野。此外,GPCRs及其配体与感染性疾病的发展之间密切相关,进一步研究可为疾病治疗提供重要的药理靶点。
利益冲突:所有作者均声明不存在利益冲突。
-
图 1 T细胞中GPCRs的功能
下载:
全尺寸图片
图 2 GPCRs在T细胞中的作用
下载:
全尺寸图片
-
[1] Audet M, Bouvier M. Restructuring G-protein- coupled receptor activation[J]. Cell, 2012, 151(1): 14-23. doi: 10.1016/j.cell.2012.09.003 [2] Yang LK, Hou ZS, Tao YX. Biased signaling in naturally occurring mutations of G protein-coupled receptors associated with diverse human diseases[J]. Biochim Biophys Acta Mol Basis Dis, 2021, 1867(1): 165973. doi: 10.1016/j.bbadis.2020.165973 [3] Hilger D, Masureel M, Kobilka BK. Structure and dynamics of GPCR signaling complexes[J]. Nat Struct Mol Biol, 2018, 25(1): 4-12. doi: 10.1038/s41594-017-0011-7 [4] Katritch V, Cherezov V, Stevens RC. Structure-function of the G protein-coupled receptor superfamily[J]. Annu Rev Pharmacol Toxicol, 2013, 53: 531-556. doi: 10.1146/annurev-pharmtox-032112-135923 [5] Dwivedi NV, Datta S, El-Kersh K, et al. GPCRs and fibroblast heterogeneity in fibroblast-associated diseases[J]. FASEB J, 2023, 37(8): e23101. doi: 10.1096/fj.202301091 [6] Hauser AS, Chavali S, Masuho I, et al. Pharmacogenomics of GPCR drug targets[J]. Cell, 2018, 172(1/2): 41-54. e19. [7] Hauser AS, Attwood MM, Rask-Andersen M, et al. Trends in GPCR drug discovery: new agents, targets and indications[J]. Nat Rev Drug Discov, 2017, 16(12): 829-842. doi: 10.1038/nrd.2017.178 [8] Chan HCS, Li Y, Dahoun T, et al. New binding sites, new opportunities for GPCR drug discovery[J]. Trends Biochem Sci, 2019, 44(4): 312-330. doi: 10.1016/j.tibs.2018.11.011 [9] Yang DH, Zhou QT, Labroska V, et al. G protein-coupled receptors: structure- and function-based drug discovery[J]. Signal Transduct Target Ther, 2021, 6(1): 7. doi: 10.1038/s41392-020-00435-w [10] Kumar BV, Connors TJ, Farber DL. Human T cell development, localization, and function throughout life[J]. Immunity, 2018, 48(2): 202-213. [11] Cosenza M, Sacchi S, Pozzi S. Cytokine release syndrome associated with T-cell-based therapies for hematological malignancies: pathophysiology, clinical presentation, and treatment[J]. Int J Mol Sci, 2021, 22(14): 7652. doi: 10.3390/ijms22147652 [12] Seder RA, Ahmed R. Similarities and differences in CD4+ and CD8+ effector and memory T cell generation[J]. Nat Immunol, 2003, 4(9): 835-842. doi: 10.1038/ni969 [13] Rosenbaum DM, Rasmussen SGF, Kobilka BK. The structure and function of G-protein-coupled receptors[J]. Nature, 2009, 459(7245): 356-363. doi: 10.1038/nature08144 [14] Syrovatkina V, Alegre KO, Dey R, et al. Regulation, signaling, and physiological functions of G-Proteins[J]. J Mol Biol, 2016, 428(19): 3850-3868. doi: 10.1016/j.jmb.2016.08.002 [15] Alkhatib G. The biology of CCR5 and CXCR4[J]. Curr Opin HIV AIDS, 2009, 4(2): 96-103. doi: 10.1097/COH.0b013e328324bbec [16] Hoft SG, Sallin MA, Kauffman KD, et al. The rate of CD4+T cell entry into the lungs during Mycobacterium tuberculosis infection is determined by partial and opposing effects of multiple chemokine receptors[J]. Infect Immun, 2019, 87(6): e00841-18. [17] Slight SR, Rangel-Moreno J, Gopal R, et al. CXCR5+T hel-per cells mediate protective immunity against tuberculosis[J]. J Clin Invest, 2013, 123(2): 712-726. [18] Harlin H, Meng YR, Peterson AC, et al. Chemokine expre-ssion in melanoma metastases associated with CD8+T-cell recruitment[J]. Cancer Res, 2009, 69(7): 3077-3085. doi: 10.1158/0008-5472.CAN-08-2281 [19] Tan MCB, Goedegebuure PS, Belt BA, et al. Disruption of CCR5-dependent homing of regulatory T cells inhibits tumor growth in a murine model of pancreatic cancer[J]. J Immunol, 2009, 182(3): 1746-1755. doi: 10.4049/jimmunol.182.3.1746 [20] Tavares LP, Garcia CC, Gonçalves APF, et al. ACKR2 contributes to pulmonary dysfunction by shaping CCL5: CCR5-dependent recruitment of lymphocytes during influenza A infection in mice[J]. Am J Physiol Lung Cell Mol Physiol, 2020, 318(4): L655-L670. doi: 10.1152/ajplung.00134.2019 [21] Kohlmeier JE, Miller SC, Smith J, et al. The chemokine receptor CCR5 plays a key role in the early memory CD8+T cell response to respiratory virus infections[J]. Immunity, 2008, 29(1): 101-113. doi: 10.1016/j.immuni.2008.05.011 [22] Contento RL, Molon B, Boularan C, et al. CXCR4-CCR5: a couple modulating T cell functions[J]. Proc Natl Acad Sci U S A, 2008, 105(29): 10101-10106. doi: 10.1073/pnas.0804286105 [23] Peng YM, van de Garde MDB, Cheng KF, et al. Specific expression of GPR56 by human cytotoxic lymphocytes[J]. J Leukoc Biol, 2011, 90(4): 735-740. doi: 10.1189/jlb.0211092 [24] Liu C, Liu TC, Hu YZ, et al. G protein-coupled receptor 56 characterizes CTLs and reflects the progression of lung cancer patients[J]. J Immunol, 2023, 211(4): 683-692. doi: 10.4049/jimmunol.2101048 [25] Kuo PT, Zeng Z, Salim N, et al. The role of CXCR3 and its chemokine ligands in skin disease and cancer[J]. Front Med (Lausanne), 2018, 5: 271. [26] Karin N. CXCR3 ligands in cancer and autoimmunity, chemoattraction of effector T cells, and beyond[J]. Front Immunol, 2020, 11: 976. doi: 10.3389/fimmu.2020.00976 [27] Li CX, Ling CC, Shao Y, et al. CXCL10/CXCR3 signaling mobilized-regulatory T cells promote liver tumor recurrence after transplantation[J]. J Hepatol, 2016, 65(5): 944-952. doi: 10.1016/j.jhep.2016.05.032 [28] Kuo P, Tuong ZK, Teoh SM, et al. HPV16E7-induced hyperplasia promotes CXCL9/10 expression and induces CXCR3+T-cell migration to skin[J]. J Invest Dermatol, 2018, 138(6): 1348-1359. doi: 10.1016/j.jid.2017.12.021 [29] Karin N, Wildbaum G, Thelen M. Biased signaling pathways via CXCR3 control the development and function of CD4+T cell subsets[J]. J Leukoc Biol, 2016, 99(6): 857-862. doi: 10.1189/jlb.2MR0915-441R [30] Groom JR. Regulators of T-cell fate: Integration of cell migration, differentiation and function[J]. Immunol Rev, 2019, 289(1): 101-114. doi: 10.1111/imr.12742 [31] Yoshie O. CCR4 as a therapeutic target for cancer immunotherapy[J]. Cancers (Basel), 2021, 13(21): 5542. doi: 10.3390/cancers13215542 [32] Zengarini C, Guglielmo A, Mussi M, et al. A narrative review of the state of the art of CCR4-based therapies in cutaneous T-cell lymphomas: focus on mogamulizumab and future treatments[J]. Antibodies (Basel), 2024, 13(2): 32. doi: 10.3390/antib13020032 [33] 刘彤, 银维谋, 蓝娇, 等. 哮喘小鼠CCR4/TARC信号轴对Treg/Th17细胞平衡的调控研究[J]. 实用医学杂志, 2020, 36(19): 2621-2624, 2629. Liu T, Yin WM, Lan J, et al. Regulation of TCR/Th17 cell infiltration into lung by CCR4/TARC signal axis in mouse asthma model[J]. The Journal of Practical Medicine, 2020, 36(19): 2621-2624, 2629. [34] Bogacka J, Pawlik K, Ciapała K, et al. CC chemokine receptor 4 (CCR4) as a possible new target for therapy[J]. Int J Mol Sci, 2022, 23(24): 15638. doi: 10.3390/ijms232415638 [35] Litvinov IV, Tetzlaff MT, Thibault P, et al. Gene expression analysis in cutaneous T-cell lymphomas (CTCL) highlights disease heterogeneity and potential diagnostic and prognostic indicators[J]. Oncoimmunology, 2017, 6(5): e1306618. doi: 10.1080/2162402X.2017.1306618 [36] Krejsgaard T, Lindahl LM, Mongan NP, et al. Malignant inflammation in cutaneous T-cell lymphoma-a hostile takeover[J]. Semin Immunopathol, 2017, 39(3): 269-282. doi: 10.1007/s00281-016-0594-9 [37] Guglielmo A, Zengarini C, Agostinelli C, et al. The role of cytokines in cutaneous T cell lymphoma: a focus on the state of the art and possible therapeutic targets[J]. Cells, 2024, 13(7): 584. doi: 10.3390/cells13070584 [38] Honjo A, Ogawa H, Azuma M, et al. Targeted reduction of CCR4+ cells is sufficient to suppress allergic airway inflammation[J]. Respir Investig, 2013, 51(4): 241-249. doi: 10.1016/j.resinv.2013.04.007 [39] 杨心怡. CCR4及Treg细胞与变应性鼻炎发病机制相关性的研究[D]. 大连: 大连医科大学, 2020. Yang XY. Correlation between CCR4 and Treg cells with the pathogenesis of allergic rhinitis[D]. Dalian: Dalian Medical University, 2020. [40] Alrumaihi F. The multi-functional roles of CCR7 in human immunology and as a promising therapeutic target for cancer therapeutics[J]. Front Mol Biosci, 2022, 9: 834149. doi: 10.3389/fmolb.2022.834149 [41] Burgstahler R, Kempkes B, Steube K, et al. Expression of the chemokine receptor BLR2/EBI1 is specifically transactiva-ted by Epstein-Barr virus nuclear antigen 2[J]. Biochem Biophys Res Commun, 1995, 215(2): 737-743. doi: 10.1006/bbrc.1995.2525 [42] Förster R, Schubel A, Breitfeld D, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs[J]. Cell, 1999, 99(1): 23-33. doi: 10.1016/S0092-8674(00)80059-8 [43] Kondo K, Ohigashi I, Takahama Y. Thymus machinery for T-cell selection[J]. Int Immunol, 2019, 31(3): 119-125. doi: 10.1093/intimm/dxy081 [44] Roberts EW, Broz ML, Binnewies M, et al. Critical role for CD103+/CD141+ dendritic cells bearing CCR7 for tumor antigen trafficking and priming of T cell immunity in melanoma[J]. Cancer Cell, 2016, 30(2): 324-336. doi: 10.1016/j.ccell.2016.06.003 [45] Sharma N, Benechet AP, Lefrançois L, et al. CD8 T cells enter the splenic T cell zones independently of CCR7, but the subsequent expansion and trafficking patterns of effector T cells after infection are dysregulated in the absence of CCR7 migratory cues[J]. J Immunol, 2015, 195(11): 5227-5236. doi: 10.4049/jimmunol.1500993 [46] Castriconi R, Carrega P, Dondero A, et al. Molecular mechanisms directing migration and retention of natural killer cells in human tissues[J]. Front Immunol, 2018, 9: 2324. doi: 10.3389/fimmu.2018.02324 [47] 秦齐雨. SHIP-1调控T细胞迁移及其在单纯疱疹病毒性角膜炎中的作用[D]. 杭州: 浙江大学, 2020. Qin QY. SHIP-1 mediated migration of CD4+T cells and their effects on herpes simplex keratitis[D]. Hangzhou: Zhejiang University, 2020. [48] Lv JW, Li L, Li W, et al. Role of the chemokine receptors CXCR3, CXCR4 and CCR7 in the intramuscular recruitment of plasmacytoid dendritic cells in dermatomyositis[J]. J Neuroimmunol, 2018, 319: 142-148. doi: 10.1016/j.jneuroim.2018.01.008 [49] 吴春玲. 次级淋巴组织趋化因子及其受体CCL21/CCR7在原发干燥综合征T淋巴细胞移行浸润中的作用及其相关机制的研究[D]. 沈阳: 中国医科大学, 2018. Wu CL. The role and mechanism of CCL21/CCR7 on T lymphocyte migration and infiltration in primary Sjögren's syndrome[D]. Shenyang: China Medical University, 2018. [50] Junger WG. Immune cell regulation by autocrine purinergic signalling[J]. Nat Rev Immunol, 2011, 11(3): 201-212. doi: 10.1038/nri2938 [51] Li RM, Tan BH, Yan Y, et al. Extracellular UDP and P2Y6 function as a danger signal to protect mice from vesicular stomatitis virus infection through an increase in IFN-β production[J]. J Immunol, 2014, 193(9): 4515-4526. doi: 10.4049/jimmunol.1301930 [52] Zhang Z, Wang ZQ, Ren H, et al. P2Y6 agonist uridine 5'-diphosphate promotes host defense against bacterial infection via monocyte chemoattractant protein-1-mediated monocytes/ macrophages recruitment[J]. J Immunol, 2011, 186(9): 5376-5387. doi: 10.4049/jimmunol.1002946 [53] Marcet B, Horckmans M, Libert F, et al. Extracellular nucleo- tides regulate CCL20 release from human primary airway epithelial cells, monocytes and monocyte-derived dendritic cells[J]. J Cell Physiol, 2007, 211(3): 716-727. doi: 10.1002/jcp.20979 [54] Giannattasio G, Ohta S, Boyce JR, et al. The purinergic G protein-coupled receptor 6 inhibits effector T cell activation in allergic pulmonary inflammation[J]. J Immunol, 2011, 187(3): 1486-1495. doi: 10.4049/jimmunol.1003669 [55] Osmers I, Smith SS, Parks BW, et al. Deletion of the G2A receptor fails to attenuate experimental autoimmune encephalomyelitis[J]. J Neuroimmunol, 2009, 207(1-2): 18-23. doi: 10.1016/j.jneuroim.2008.11.008 [56] Hasegawa H, Lei J, Matsumoto T, et al. Lysophosphatidylcholine enhances the suppressive function of human naturally occurring regulatory T cells through TGF-β production[J]. Biochem Biophys Res Commun, 2011, 415(3): 526-531. doi: 10.1016/j.bbrc.2011.10.119 [57] Yang LV, Radu CG, Wang L, et al. Gi-independent macrophage chemotaxis to lysophosphatidylcholine via the immunoregulatory GPCR G2A[J]. Blood, 2005, 105(3): 1127-1134. doi: 10.1182/blood-2004-05-1916 [58] Radu CG, Yang LV, Riedinger M, et al. T cell chemotaxis to lysophosphatidylcholine through the G2A receptor[J]. Proc Natl Acad Sci U S A, 2004, 101(1): 245-250. [59] Cotte AK, Aires V, Fredon M, et al. Lysophosphatidylcholine acyltransferase 2-mediated lipid droplet production su-pports colorectal cancer chemoresistance[J]. Nat Commun, 2018, 9(1): 322. doi: 10.1038/s41467-017-02732-5 [60] Peter C, Waibel M, Radu CG, et al. Migration to apoptotic "find-me" signals is mediated via the phagocyte receptor G2A[J]. J Biol Chem, 2008, 283(9): 5296-5305. doi: 10.1074/jbc.M706586200 [61] Kalinski P. Regulation of immune responses by prostaglandin E2[J]. J Immunol, 2012, 188(1): 21-28. doi: 10.4049/jimmunol.1101029 [62] Li H, Chen HY, Liu WX, et al. Prostaglandin E2 restrains human Treg cell differentiation via E prostanoid receptor 2-protein kinase A signaling[J]. Immunol Lett, 2017, 191: 63-72. doi: 10.1016/j.imlet.2017.09.009 [63] Nataraj C, Thomas DW, Tilley SL, et al. Receptors for prostaglandin E2 that regulate cellular immune responses in the mouse[J]. J Clin Invest, 2001, 108(8): 1229-1235. doi: 10.1172/JCI200113640 [64] Boniface K, Bak-Jensen KS, Li Y, et al. Prostaglandin E2 regulates Th17 cell differentiation and function through cyclic AMP and EP2/EP4 receptor signaling[J]. J Exp Med, 2009, 206(3): 535-548. doi: 10.1084/jem.20082293 [65] Sreeramkumar V, Hons M, Punzón C, et al. Efficient T-cell priming and activation requires signaling through prostaglandin E2 (EP) receptors[J]. Immunol Cell Biol, 2016, 94(1): 39-51. doi: 10.1038/icb.2015.62 [66] Snyder JP, Amiel E. Regulation of dendritic cell immune function and metabolism by cellular nutrient sensor mammalian target of rapamycin (mTOR)[J]. Front Immunol, 2019, 9: 3145. doi: 10.3389/fimmu.2018.03145 [67] Pompura SL, Dominguez-Villar M. The PI3K/AKT signaling pathway in regulatory T-cell development, stability, and function[J]. J Leukoc Biol, 2018, 103(6): 1065-1076. doi: 10.1002/JLB.2MIR0817-349R [68] Dinkel BA, Kremer KN, Rollins MR, et al. GRK2 mediates TCR-induced transactivation of CXCR4 and TCR-CXCR4 complex formation that drives PI3Kγ/PREX1 signaling and T cell cytokine secretion[J]. J Biol Chem, 2018, 293(36): 14022-14039. doi: 10.1074/jbc.RA118.003097 [69] Kammala AK, Yang CC, Panettieri RA, et al. G protein-coupled receptor kinase 2 (GRK2) regulates T cell response in a murine model of house dust mite-induced asthma[J]. Front Allergy, 2021, 2: 656886. doi: 10.3389/falgy.2021.656886 [70] Wang QT, Wang LS, Wu L, et al. Paroxetine alleviates T lymphocyte activation and infiltration to joints of collagen-induced arthritis[J]. Sci Rep, 2017, 7: 45364. doi: 10.1038/srep45364 [71] 金超. 羟氯喹抑制T细胞中GRK2与PI3K相互作用抗自身免疫性肝炎[D]. 合肥: 安徽医科大学, 2023. Jin C. Hydroxychloroquine attenuates autoimmune hepatitis by suppressing the interaction of GRK2 with PI3K in T lymphocytes[D]. Hefei: Anhui Medical University, 2023. [72] Cekic C, Day YJ, Sag D, et al. Myeloid expression of adenosine A2A receptor suppresses T and NK cell responses in the solid tumor microenvironment[J]. Cancer Res, 2014, 74(24): 7250-7259. [73] Ohta A, Sitkovsky M. Role of G-protein-coupled adenosine receptors in downregulation of inflammation and protection from tissue damage[J]. Nature, 2001, 414(6866): 916-920. [74] Kjaergaard J, Hatfield S, Jones G, et al. A2A adenosine receptor gene deletion or synthetic A2A antagonist liberate tumor-reactive CD8+T cells from tumor-induced immunosu-ppression[J]. J Immunol, 2018, 201(2): 782-791. [75] Leone RD, Sun IM, Oh MH, et al. Inhibition of the adenosine A2a receptor modulates expression of T cell coinhibitory receptors and improves effector function for enhanced checkpoint blockade and ACT in murine cancer models[J]. Cancer Immunol Immunother, 2018, 67(8): 1271-1284. [76] Mediavilla-Varela M, Castro J, Chiappori A, et al. A novel antagonist of the immune checkpoint protein adenosine A2a receptor restores tumor-infiltrating lymphocyte activity in the context of the tumor microenvironment[J]. Neoplasia, 2017, 19(7): 530-536. [77] Ma SR, Deng WW, Liu JF, et al. Blockade of adenosine A2A receptor enhances CD8+T cells response and decreases regulatory T cells in head and neck squamous cell carcinoma[J]. Mol Cancer, 2017, 16(1): 99. [78] Mastelic-Gavillet B, Navarro Rodrigo B, Décombaz L, et al. Adenosine mediates functional and metabolic suppression of peripheral and tumor-infiltrating CD8+T cells[J]. J Immunother Cancer, 2019, 7(1): 257. [79] Revankar CM, Cimino DF, Sklar LA, et al. A transmembrane intracellular estrogen receptor mediates rapid cell signaling[J]. Science, 2005, 307(5715): 1625-1630. [80] Chakrabarti S, Davidge ST. G-protein coupled receptor 30 (GPR30): a novel regulator of endothelial inflammation[J]. PLoS One, 2012, 7(12): e52357. [81] Prossnitz ER, Barton M. The G-protein-coupled estrogen receptor GPER in health and disease[J]. Nat Rev Endocrinol, 2011, 7(12): 715-726. [82] Wang CH, Dehghani B, Li YX, et al. Membrane estrogen receptor regulates experimental autoimmune encephalomyelitis through up-regulation of programmed death 1[J]. J Immunol, 2009, 182(5): 3294-3303. [83] Bernardi AI, Andersson A, Stubelius A, et al. Selective estrogen receptor modulators in T cell development and T cell dependent inflammation[J]. Immunobiology, 2015, 220(10): 1122-1128. [84] Wang CH, Dehghani B, Magrisso IJ, et al. GPR30 contri-butes to estrogen-induced thymic atrophy[J]. Mol Endocrinol, 2008, 22(3): 636-648. [85] Fisher J, Card G, Soong L. Neuroinflammation associated with scrub typhus and spotted fever group rickettsioses[J]. PLoS Negl Trop Dis, 2020, 14(10): e0008675. [86] Mamtani M, Mummidi S, Ramsuran V, et al. Influence of variations in CCL3L1 and CCR5 on tuberculosis in a northwestern Colombian population[J]. J Infect Dis, 2011, 203(11): 1590-1594. [87] Shanmugasundaram U, Bucsan AN, Ganatra SR, et al. Pulmonary Mycobacterium tuberculosis control associates with CXCR3- and CCR6-expressing antigen-specific Th1 and Th17 cell recruitment[J]. JCI insight, 2020, 5(14): 137858. [88] Park HS, Choi S, Back YW, et al. Mycobacterium tuberculosis RpfE-induced prostaglandin E2 in dendritic cells induces Th1/Th17 cell differentiation[J]. Int J Mol Sci, 2021, 22(14): 7535. [89] Mergia Terefe E, Catalan Opulencia MJ, Rakhshani A, et al. Roles of CCR10/CCL27-CCL28 axis in tumour development: mechanisms, diagnostic and therapeutic approaches, and perspectives[J]. Expert Rev Mol Med, 2022, 24: e37. [90] Ohue Y, Nishikawa H. Regulatory T (Treg) cells in cancer: can Treg cells be a new therapeutic target?[J]. Cancer Sci, 2019, 110(7): 2080-2089. [91] Fujita H. The role of IL-22 and Th22 cells in human skin di-seases[J]. J Dermatol Sci, 2013, 72(1): 3-8. [92] Xiong J, Dai YT, Wang WF, et al. GPCR signaling contri-butes to immune characteristics of microenvironment and process of EBV-induced lymphomagenesis[J]. Sci Bull (Beijing), 2023, 68(21): 2607-2619. [93] 张娜. 病毒感染过程中关键GPCR的筛选及其功能和机制探究[D]. 上海: 华东师范大学, 2017. Zhang N. Identification and mechanism study of key GPCRs during viral infection[D]. Shanghai: East China Normal University, 2017.
